Химия закиси азота - свойства N₂O
Подробно о структуре и реакции закиси азота: формула, получение в лаборатории и промышленности, устойчивость и взаимодействия.
Search
Have an existing account?
Sign In






Сравнительный анализ медицинской и немедицинской закиси азота
Сравнительный анализ медицинской и немедицинской закиси азота Закись азота, известная также как «газ веселья», представляет собой химическое соединение с формулой…




Эксперты раскрывают основные отличия между медицинскими и пищевыми баллонами
Эксперты раскрывают основные отличия между медицинскими и пищевыми баллонами В современном мире баллоны используются в…

Закись азота: новое средство для изучения сознания
Закись азота: новое средство для изучения сознания Закись азота, более известная как "газ смеха", уже…


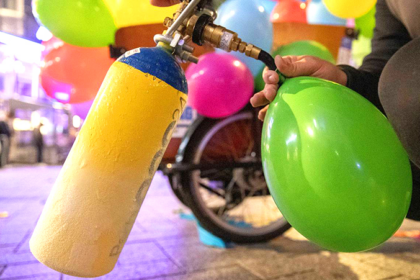
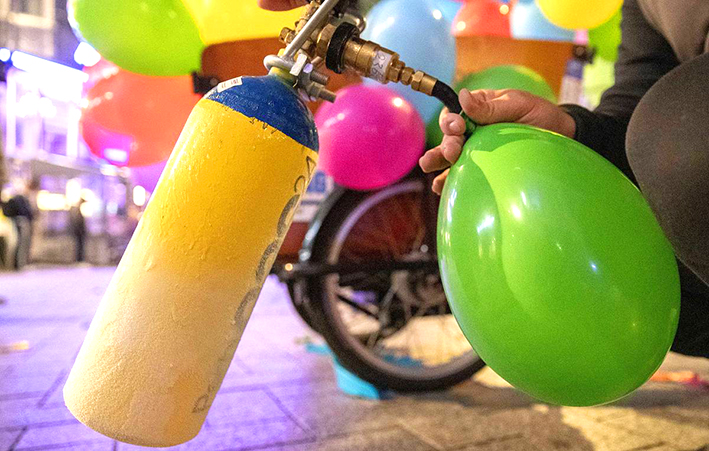
В чем различия между закисью азота, пропаном и бутаном?
В чем различия между закисью азота, пропаном и бутаном? Закись азота, пропан и бутан — это три различных вещества, которые…








